
Colour has been reconstructed in birds from the Jurassic, Cretaceous and Cenozoic – it seems that their melanosomes are equally well preserved through all geological time. There are not yet enough detailed studies to enable palaeontologists to explore how colour, and assumed functions and behaviours associated with those colours and patterns, might have evolved. This is for the future! This page reviews briefly some of the recent studies, in age sequence.
Jurassic of Germany

One of the first birds to be studied for original colour was Archaeopteryx from the Solnhofen beds of southern Germany. The feather sampled was an isolated specimen found in 1860, and it indicated black feathers based on numerous, closely packed, elongate eumelanosomes (Carney et al. 2012). The study led to many reconstructions of the ‘first bird’ Archaeopteryx as all-black, quite in contrast to more traditional reconstructions where its plumage had been shown as a mix of colours, although those earlier reconstructions were based on no evidence.
Subsequently, some doubt was cast on whether the isolated feather really came from Archaeopteryx or not; Kaye et al. (2019) suggested the feather could be a covert or contour feather of Archaeopteryx, as Carney et al. (2012) had indicated, or, they said, ‘the possibility remains that it stems from a different feathered dinosaur that lived in the Solnhofen Archipelago.’
[Right] The isolated fossil feather from the Late Jurassic Solnhofen beds (MB.Av.100). Scale bar: 5 mm.
Cretaceous of Brazil
The first report of melanosomes in fossils was by Jakob Vinther and colleagues from Yale University (Vinther et al. 2008). He noted that the dark stripes of a fossil feather from the Cretaceous of Brazil contained tiny sausage-shaped structures, but the pale stripes showed nothing but rock. He argued these structures were not bacteria, as had been thought, but melanosomes, the structures that contain the pigment melanin in modern bird feathers.
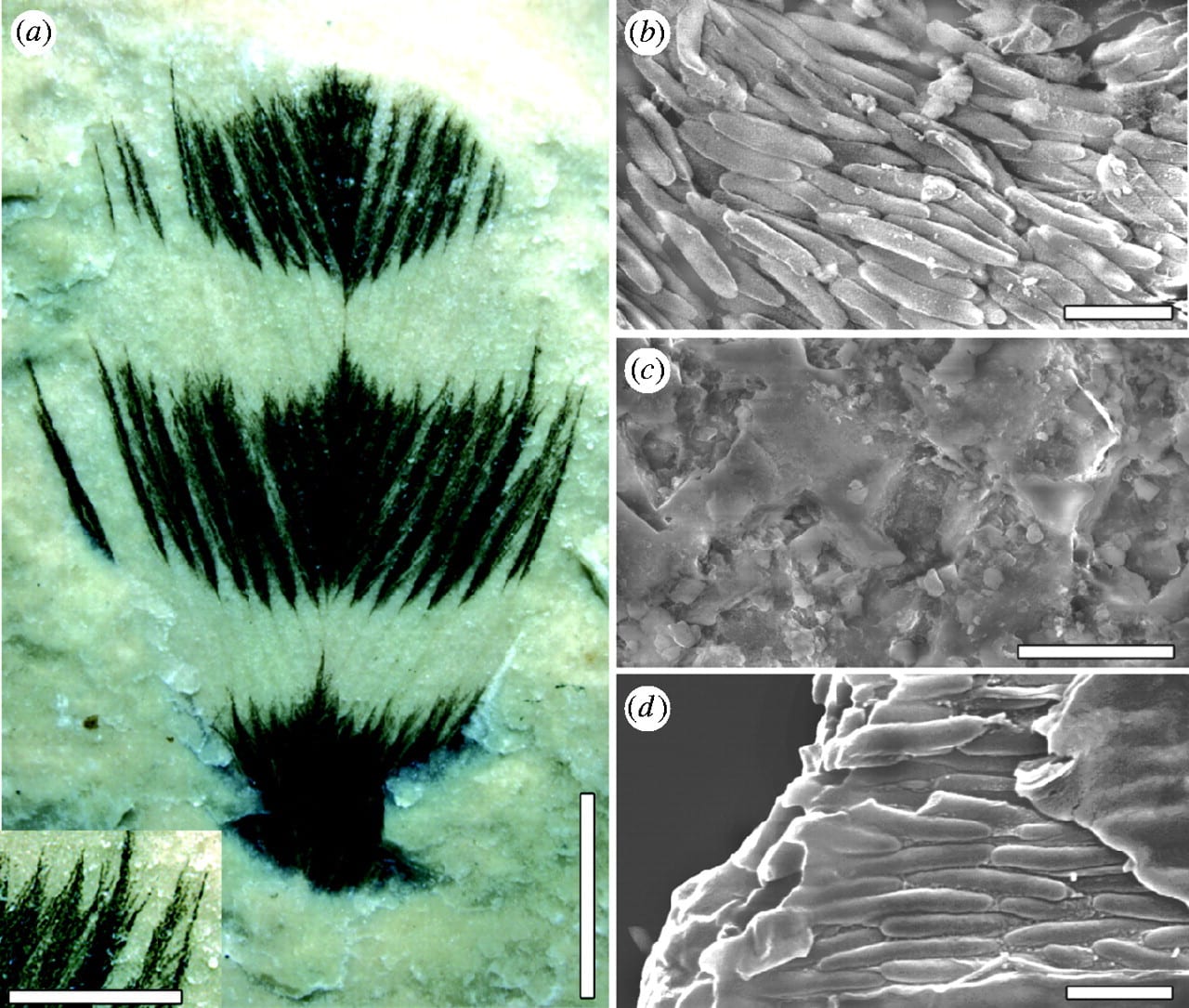 Figure 1. Cretaceous feather ultrastructure compared with that in a living bird. (a) Feather from the Crato Formation, Early Cretaceous, Brazil showing colour bands; margins of colour bands are similar to those found in living birds and barbules are clearly preserved. (b) Dark bands, composed of aligned eumelanosomes, contrast with (c) light areas that reveal only the rock matrix. (d) A broken barbule from a modern Red-winged Blackbird (Agelaius phoeniceus) reveals eumelanosomes aligned along the barbule enclosed in a keratin matrix. Scale bars, (a) 3 mm, insert 1 mm; (b) 1 µm; (c) 10 µm; (d ) 1 µm. [Image from Jakob Vinther.]
Figure 1. Cretaceous feather ultrastructure compared with that in a living bird. (a) Feather from the Crato Formation, Early Cretaceous, Brazil showing colour bands; margins of colour bands are similar to those found in living birds and barbules are clearly preserved. (b) Dark bands, composed of aligned eumelanosomes, contrast with (c) light areas that reveal only the rock matrix. (d) A broken barbule from a modern Red-winged Blackbird (Agelaius phoeniceus) reveals eumelanosomes aligned along the barbule enclosed in a keratin matrix. Scale bars, (a) 3 mm, insert 1 mm; (b) 1 µm; (c) 10 µm; (d ) 1 µm. [Image from Jakob Vinther.]
The feather came from the Crato Formation (mid Cretaceous, c. 125 million years ago; Fig. 1a), and it shows closely packed eumelanosomes in the black stripes (Fig. 1b), but just the rock matrix, and no melanosomes on the white stripes (Fig. 1c). The sharp boundary between the black and white stripe is unlikely to reflect any later damage to the feather, and this is confirmed by close microscopic examination: the black band shows more relief, produced the strenghtening properties of the melanosomes themselves, and chemical analysis shows the presence of carbon in the black stripes, perhaps a residue of the original eumelanin, but not in the white stripes. Finally, the sharp contact, and the V shape of the stripe matches patterns seen in modern bird feathers.
Cretaceous of China
Most work on fossil feathers has focused on the Cretaceous of China. Since the first discovery of fossil birds such as Confuciusornis in the Jehol Group of north-east China, palaeontologists have investigated their feathers. In 2010, Zhang et al. (2010) were first to identify both eumelanosomes and phaeomelanosomes in bird and dinosaur feathers (Fig. 2).
Figure 2. Melanosomes in feathers of the bird Confuciusornis (IVPP V13171). a, Optical photograph. b, Strongly aligned, closely spaced, eumelanosomes preserved as solid bodies. c, Mouldic (that is, preserved as moulds) eumelanosomes (at arrow) a short distance above a layer in which the eumelanosomes are preserved as aligned solid bodies. d, Area (at arrow) comprising more widely spaced mouldic phaeomelanosomes surrounded by less distinct, aligned eumelanosomes (top of image). e, Gradational boundary between areas dominated by eumelanosomes (longer arrows) and phaeomelanosomes (shorter arrows), both preserved as solid bodies. Scale bars: a, 50 mm; b-e, 2 µm. [Image by Stuart Kearns.]
Feathers of Confuciusornis (above), and other Jehol birds, show both eumelanosomes and phaeomelanosomes. In the Confuciusornis feathers (Fig. 2), preservation of the degraded keratinous matrix, presumably as an organic remain, occurs locally in some of the Jehol feathers, most obviously where the fossil bodies are exposed as moulds (Fig. 2c, d); the fossil bodies are, like melanosomes, clearly embedded within this matrix (Fig. 2e), and are not a superficial coating.
 In Confuciusornis, eumelanosomes often occur closely packed, strongly aligned and forming a discrete layer (Figs. 2b, d), as in other fossil birds (Vinther et al. 2008), and the arrays of melanosomes in extant birds (Durrer 1986). In isolated feathers, the eumelanosomes occur in well-defined areas (each c. 20 mm long and 6-8 mm wide) that are separated by narrow (c. 2 mm wide), anastomosing, ridges of degraded feather (Fig. 1b, c). The eumelanosomes are strongly aligned, parallel to the long axis of each area (Fig. 2c). Phaeomelanosomes may occur as areas surrounded by eumelanosomes, as in Confuciusornis (Fig. 2d).
In Confuciusornis, eumelanosomes often occur closely packed, strongly aligned and forming a discrete layer (Figs. 2b, d), as in other fossil birds (Vinther et al. 2008), and the arrays of melanosomes in extant birds (Durrer 1986). In isolated feathers, the eumelanosomes occur in well-defined areas (each c. 20 mm long and 6-8 mm wide) that are separated by narrow (c. 2 mm wide), anastomosing, ridges of degraded feather (Fig. 1b, c). The eumelanosomes are strongly aligned, parallel to the long axis of each area (Fig. 2c). Phaeomelanosomes may occur as areas surrounded by eumelanosomes, as in Confuciusornis (Fig. 2d).
[Left] Figure 3. Confuciusornis reconstruction by Velizar Simeonovski (Creative Commons).
In other work on Cretaceous bird feathers, Wogelius et al. (2011) reconstructed the coloration of Confuciusornis using trace metal chemistry of fossil melanin. On a related theme, Edwards et al. (2016) showed chemical evidence of phaeomelanin in modern feathers using Zn mapping – a potential proxy for phaeomelanin in fossils, which thus far has proved elusive.
Eocene of Denmark and Germany
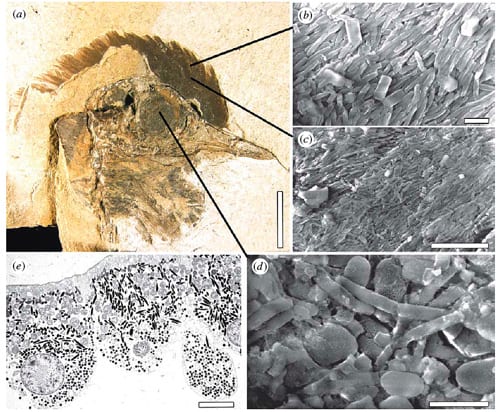 Figure 4. (a) Skull of undescribed bird from the Fur Formation, Early Eocene, Denmark, preserving feathers and the eye as an organic film. (b, c) Details of the feather region showing aligned eumelanosomes. (d) Detail of the eye showing elongate and oblate eumelanosomes. (e) TEM of a section through the retina of a Whip-poor-will (Caprimulgus vociferus, Caprimulgidae). Scale bars, (a) 10 mm; (b) 1 µm; (c) 5 µm; (d) 1 µm; (e) 5 µm. [Image by Jakob Vinther.]
Figure 4. (a) Skull of undescribed bird from the Fur Formation, Early Eocene, Denmark, preserving feathers and the eye as an organic film. (b, c) Details of the feather region showing aligned eumelanosomes. (d) Detail of the eye showing elongate and oblate eumelanosomes. (e) TEM of a section through the retina of a Whip-poor-will (Caprimulgus vociferus, Caprimulgidae). Scale bars, (a) 10 mm; (b) 1 µm; (c) 5 µm; (d) 1 µm; (e) 5 µm. [Image by Jakob Vinther.]
Vinther et al. (2008) described melanosomes from an unnamed bird from the early Eocene Fur Formation, some 55 million years old. They examined the head region (Fig. 4a), and identified eumelanosomes in the head feathers (Fig. 4b, c) and in black material from the retina of the eye (Fig. 4d).
Vinther et al. (2008) also identified eumelanosomes from the Messel locality in Germany, middle Eocene (47 Myr) old, world famous for the exceptional preservation of feathers, hair, and other soft tissues in a wide range of animals. In a second study, Vinther et al. (2010) investigated the Messel feathers further, and they reported arrays of aligned rod-shaped eumelanosomes (Fig. 5). In some, they noted a continuous external layer of closely packed melanosomes enclosing loosely aligned melanosomes, similar to the single thin-film nanostructure that generates an iridescent, structurally coloured sheen on the surface of black feathers in many lineages of living birds. This was the first evidence of preservation of a colour-producing nanostructure in a fossil feather.
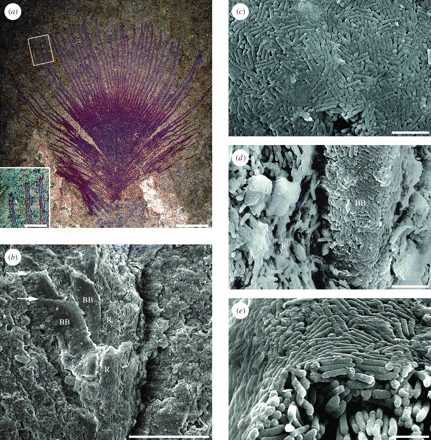
Figure 5. Open pennaceous contour feather from the Eocene Messel Oil Shale, showing evidence of original structural colour. (a) The part; the inset shows a detail of the distal barb rami and reflective barbules. (b-e) SEMs of samples from the counterpart. (b) Distal part of barb ramus. (c) Surface of the barbule melanosome layer. (d) Base of a barbule and its attachment to the barb ramus; note the lack of a uniform melanosome layer on the ramus. (e) Cross-section of barbule showing thin outer layer and aligned melanosomes within. R, ramus with loose aligned melanosomes; BB, associated barbules with surface layer of melanosomes; arrows show cell boundaries. Scale bars: (a) 5 mm, inset 1 mm; (b) 50 µm; (c,d) 2 µm; (e) 1 µm. [Image by Jakob Vinther.]
Eocene of Peru
 In a study of the Eocene penguin Inkayacu from Peru, Clarke et al. (2010) report a mix of eumelanosomes and phaeomelanosomes, indicating grey and reddish-brown feather colours. In contrast, the dark black-brown colour of extant penguin feathers is generated by large, ellipsoidal melanosomes previously unknown for birds – these melanosomes in modern penguins are about the same length as eumelanosomes (c. 900 nm), but they are wider (c. 440 nm, compared to c. 300 nm). The fossil then retains the normal melanosomes seen in other birds living and fossil, but lacks the specialized melanosomes of modern penguins which must then have evolved later.
In a study of the Eocene penguin Inkayacu from Peru, Clarke et al. (2010) report a mix of eumelanosomes and phaeomelanosomes, indicating grey and reddish-brown feather colours. In contrast, the dark black-brown colour of extant penguin feathers is generated by large, ellipsoidal melanosomes previously unknown for birds – these melanosomes in modern penguins are about the same length as eumelanosomes (c. 900 nm), but they are wider (c. 440 nm, compared to c. 300 nm). The fossil then retains the normal melanosomes seen in other birds living and fossil, but lacks the specialized melanosomes of modern penguins which must then have evolved later.
[Right] Reconstruction of original feather colours in the fossil penguin Inkayacu. [Artwork by Katie Browne.]
Literature cited
- Carney, R.M., Vinther, J., Shawkey, M.D., D’Alba L., and Ackerman, J. 2012. New evidence on the colour and nature of the isolated Archaeopteryx feather. Nature Communications 3, 637.
- Clarke, J.A., Kspeka, D.T., Salas-Gismondi, R., Altamirano, A.J., Shawkey, M.D., D’Alba, L., Vinther, J., DeVries, T.J., and Baby, P. 2010. Fossil evidence for evolution of the shape and color of penguin feathers. Science 330, 954-957.
- Durrer, H. 1986. The skin of birds: Colouration. In Biology of the Integument 2, Vertebrates (eds Bereiter-Hahn, J., Matolsky, A.G. & Richards, K.S.), pp. 239-247. Springer.
- Edwards, N.P. 2016. Elemental characterisation of melanin in feathers via synchrotron X-ray imaging and absorption spectroscopy. Scientific Reports 6, art. 34002.
- Kaye, T.G., Pittman, M., Mayr, G., Schwarz, D. and Xu, X. 2019. Detection of lost calamus challenges identity of isolated Archaeopteryx feather. Scientific Reports 9, 1182.
- Vinther, J., Briggs, D. E. G., Prum, R. O. & Saranathan, V. 2008. The colour of fossil feathers. Biology Letters 4, 522-525.
- Vinther, J., Briggs, D. E. G., Clarke, J., Mayr, G. & Prum, R. O. 2010. Structural coloration in a fossil feather. Biology Letters 6, 128-131.
- Wogelius, R.A. et al. 2011. Trace metals as biomarkers for eumelanin pigment in the fossil record. Science 333, 1622–1626.
- Zhang, F., Kearns, S.L, Orr, P.J., Benton, M.J., Zhou, Z., Johnson, D., Xu, X., and Wang, X. 2010. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 463, 1075-1078. Download the paper here and the Supplementary data.